






Генетика любви: межполовой конфликт как основа сотрудничества в парах моногамных птиц
Отношения между партнерами, наполненные заботой, знаками внимания и сопереживанием, поэты называют любовью, а вот биологи — межполовой взаимосвязью, нацеленной на выживание и продолжение рода. Одни виды предпочитают брать количеством — размножаться с как можно большим числом партнеров, чтобы увеличить потомство, тем самым повысив шансы на выживание всего вида. Другие же создают моногамные пары, которые могут прекратить свое существование лишь после смерти одного из партнеров.

Многие годы ученые считали, что первый вариант значительно выгоднее, но это не совсем так. Моногамные пары, как правило, совместно воспитывают потомство, т.е. защищают его от хищников, добывают пищу и учат определенным навыкам, тогда как в полигамных отношениях все это чаще всего ложится на хрупкие плечи самок. Конечно, бывают исключения, но сегодня не о них. Биологов долгое время интересовал другой занятный момент — самцы продолжают проявлять знаки внимания в адрес самок, даже когда их пара уже сформирована и существует несколько лет. Чем обусловлено такое поведение, какая от этого выгода и какие эволюционные аспекты с этим связаны? На эти вопросы мы найдем ответы в докладе исследовательской группы. Поехали.
Основа исследования
Учитывая тематику исследования, мы не будем заострять внимание на полигамных видах птиц, а сосредоточимся на пернатых романтиках, которые влюбляются раз и навсегда.
Говоря о моногамии, стоит отметить, что существует несколько ее видов в зависимости от продолжительности: один сезон, несколько лет и на всю жизнь.
Среди птиц наиболее распространенной является сезонная моногамия. Ярким примером могут быть дикие гуси. Самки занимаются гнездом и высиживанием яиц, а самец охраной территории. На второй день после вылупления семейство отправляется к ближайшему водоему, где гусята учатся искать себе пропитание. В случае опасности, настигшей на воде, самка яростно защищает потомство, а вот самец, видимо вспомнив про важные дела, чаще всего спасается бегством. Не самые идеальные отношения, как ни крути.

Семейство диких гусей.
Если же говорить об отношениях, основой которых является постоянство, то аисты — лучшие в этом вопросе. Они создают моногамную пару на всю жизнь и даже место жительства не меняют без особой на то надобности. Одно гнездо аистов, которое может весить до 250 кг и достигать 1.5 м в диаметре, служит им много лет, если природные катаклизмы или человеческое вмешательство его не разрушат. В Чехии есть гнездо, которое было создано еще в 1864 году.

Строительные навыки аистов не нуждаются в оценке, когда видишь такие сооружения.
В отличие от диких гусей, у аистов царствует равенство обязанностей: оба партнера высиживают яйца, ищут пищу, учат потомство летать и защищают его от опасностей. Важную роль в отношениях аистов играют различного рода ритуалы: пение, танцы и т.д. Самое любопытное, что эти ритуалы проводятся не только в период формирования пары (на первом свидании), но и в течение всей совместной жизни (даже сменяя самку во время высиживания самец исполняет небольшой танец). Для нас это выглядит очень мило, романтично и абсолютно нелогично, поскольку с биологической точки зрения у такого поведения нет никакой выгоды. Ведь так? И вот тут можно плавненько приступить к рассмотрению самого исследования, которое и должно было ответить на этот вопрос.
Этологи* считают, что постоянное проявление самцами своих чувств связано с сохранением у самок репродуктивного состояния.
Этология* — наука, изучающая генетически обусловленное поведение, т.е. инстинкты.
В то же время остается неясно, почему такое поведение длится не только в период первичного спаривания, а всю жизнь, ибо самцам логичнее было бы вкладывать больше сил и энергии в потомство, а не в демонстрацию чувств к самке. К сегодняшнему дню многие исследователи считали, что интенсивность выражения привязанности к самке напрямую влияет на качество спаривания и, следовательно, потомства (т.е. число отложенных яиц).

Самец одного из видов райских птиц танцует перед самкой. Как мы видим, самец выглядит намного ярче самки.
Эта теория подтверждается наблюдениями. Самка, чей партнер неписаный красавец и первый летун на селе, вкладывает больше усилий в потомство, нежели если самец ни рыба ни мясо. Звучит забавно и смешно, но ритуалы, которые проводят самцы перед самками нацелены на демонстрацию не только красоты, но и силы. Так уж вышло, что яркое оперения, красивое пение и другие проявления внимания со стороны самцов являются для самок всего лишь когнитивными сигналами, которые она декодирует в информацию о самце.
Ученые из университетов Северной Каролины и Чикаго, чей труд мы сегодня рассматриваем, считают, что подобное поведение самцов нацелено на оптимизацию поведения самок по отношению к процессу выведения потомства.
Модель, предложенная учеными, основана на многочисленных экспериментах, которые показали, что усиление данных сигналов со стороны самцов увеличивает вклад самок в процесс продолжения рода. Существует предположение, что источником подобных стимулирующих эффектов являются перцептивные ответы, возникающие из-за свойств окружающей среды, сигналов и нервной системы как таковой. На данный момент известно порядка 100 примеров подобных «отклонений» от обычных сенсорных систем (слух, зрение и запах).
Когда самец лишний раз демонстрирует свои преимущества перед другими самцами, это может положительно повлиять на самого самца (самка его точно выберет). Но для самки это может быть минусом, так как это уменьшит будущую репродуктивную реализацию. Другими словами, мы имеем ситуацию «превзойти ожидания». Самец, который значительно лучше других самцов и постоянно проявляет знаки интереса к самке, получит желаемое — спаривание и продолжение рода, а точнее своего рода. Самка же, ожидающая от других самцов подобного поведения, но не получившая его, может оказаться в плачевной ситуации. Подобный случай именуется учеными как межполовой конфликт: демонстрация самцами себя красивых увеличивается среди популяции, а среди самок растет сопротивление этой тактике.
Данный конфликт был смоделирован с применением вычислительного подхода (нейронные сети). В полученных моделях сигнализатор (источник сигнала — самец) использует перцептивное восприятие приемника (приемник сигнала — самка), которое стимулирует сами сигналы в ущерб восприятию. В определенный момент возникает изменение восприятия сигналов в популяции самок (своего рода мутация), от чего сила сигналов источника (самца) будет сильно снижаться. Постепенное нарастание таких изменений приведет к тому, что тот или иной тип сигнала будет абсолютно неэффективен. По мере протекания таких изменений одни сигналы исчезают, утратив свою силу, но возникают новые, и процесс начинается заново.
Эта весьма закрученная система достаточно проста на практике. Представьте, что появился самец с ярким пером (одним единственным), он выделяется на фоне других, и самки отдают предпочтение именно ему. Потом появляется самец с двумя яркими перышками, потом с тремя и т.д. Но сила подобного сигнала, ввиду его нарастания и распространения, начинает пропорционально падать. И тут вдруг появляется самец, который умеет красиво петь и строить гнезда. В результате красивое оперение как сигнал перестает быть эффективным и начинает вырождаться.
Однако всегда есть исключение из правил — некоторые межполовые конфликты могут перерасти в полноценное и весьма эффективное межполовое сотрудничество.
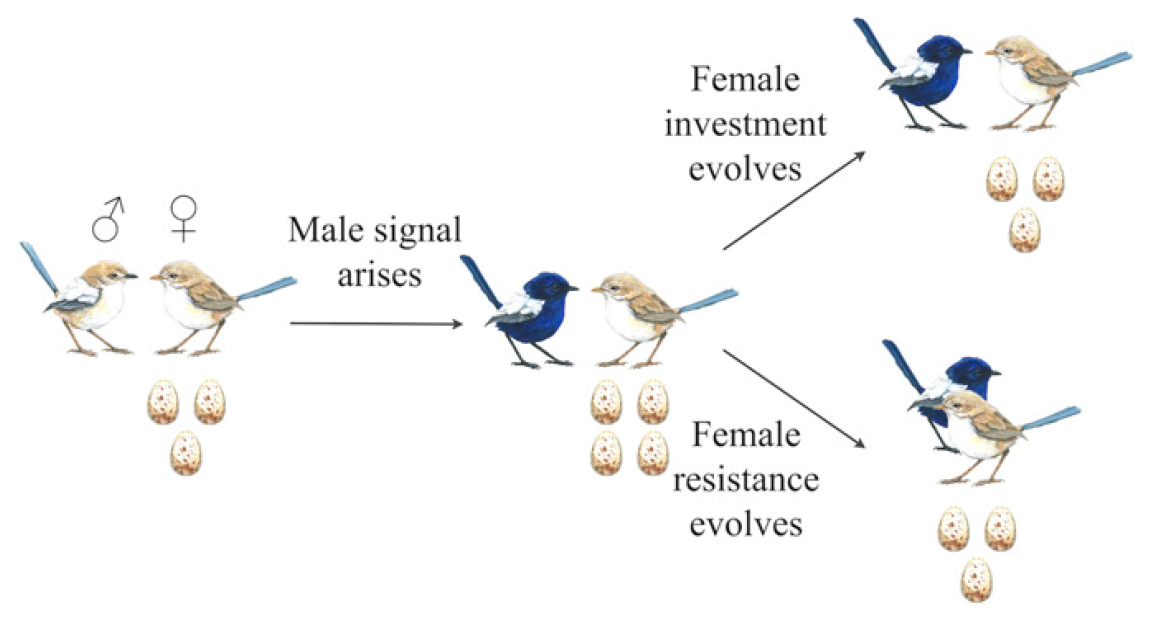
Схема возникновения межполового конфликта и межполового сотрудничества.
Суть заключается в том, что самец с более выраженным сигналом вынуждает самку откладывать не три яйца, а четыре. Для самца это хорошо — у него будет больше потомства с его генофондом. Для самки — не очень, ибо ей придется потратить больше сил на то, чтобы все потомство выжило и достигло самостоятельного возраста. Следовательно, самки начинают развиваться параллельно самцам, чтобы быть более устойчивыми к их сигналам. В результате может быть два пути: конфликт или сотрудничество.
В случае сотрудничества самки эволюционируют таким образом, чтобы откладывать 3 яйца, как и до появления более сильного сигнала со стороны самцов, но продолжать реагировать на эти сигналы. Вот тебе и женские хитрости в мире природы. Таким образом формируется не просто пара, а пара, поддерживающая друг друга на оптимальном уровне для продолжения рода с точки зрения взаимодействия сигнал-ответ.
Самцы же не могут эволюционировать обратно, грубо говоря. Их усиленные сигналы в адрес самок дают результат в виде кладки из трех яиц, т.е. не такой, как ожидалось. Однако снижение сигнала до предыдущего уровня будет также неэффективным, так как приведет к уменьшению числа яиц в кладке до двух. Получается замкнутый круг — самцы не могут уменьшить силу сигнала и не могут ее увеличить, так как самки в первом случае будут давать меньше потомства, а во втором — не реагировать.
Естественно, ни у самцов, ни у самок нет никакого злого умысла или желания поработить друг друга. Весь этот процесс протекает на генетическом уровне и нацелен исключительно во благо потомства отдельно взятой пары и благополучия вида в целом.
Результаты исследования
Используя математическое моделирование, ученые оценили условия, при которых может возникнуть межполовое сотрудничество. Количественная характеристика со средним значением zf описывает основной вклад самки в ее потомство. Изначально среднему значению разрешается развиваться до своего оптимального значения zopt, которое зависит от двух переменных: выгода от вклада (количество выживших потомков) и стоимость вклада для самок (cf). Последняя переменная оценивается после размножения, подразумевая, что часть самок выживает и может в следующем году еще раз произвести потомство, что приводит к увеличению числа поколений.
В данном исследовании будет частенько применяться несколько терминов, которые стоит немного пояснить:
- сигналы — проявление внимания со стороны самцов в адрес партнеров-самок (пение, танцы и другие ритуалы), имеющие место в сформированных парах;
- вклад / инвестиции — ответ самок на эти сигналы, проявляющиеся в виде большего числа яиц в кладке, большего времени на уход за будущим потомством и т.д.;
- респондент — самка, реагирующая на сигналы самца;
- затраты — стоимость вклад самок в потомство (время в гнезде, время на поиски пищи, состояние здоровья ввиду большего/меньшего числа яиц в кладке и т.д.).
Новые сигналы самцов и реакции самок на них моделировались посредством свободно рекомбинирующих диаллельных одно-локусных модификаторов, таким образом объединяя количественный и популяционный генетические подходы. В локусе*, который контролирует ответ самки (A), первоначально наблюдается высокая частота аллеля-респондера* (A2), соответствующая существовавшему ранее перцептивному восприятию
Локус* — местоположение определенного гена на генетической карте хромосомы.
Аллели* — различные формы одного и того же гена, расположенные в одинаковых локусах гомологичных хромосом. Аллели определяют путь развития конкретного признака.
Ген-респондер* (Rsp) — ген, функционально связанный с фактором нарушения сегрегации (ген SD), активный аллель которого (Rsp+) способен подавлять экспрессию SD.
Локус сигнала (B) изначально фиксирован для аллеля без сигнала (B1). Затем вводится аллель B2, который вызывает появление сигналов со стороны самцов.
Проявление сигналов для самцов также имеет свою цену (sm), но увеличивает вклад партнера-самки (А2) на величину α. К примеру, α может быть выражено в виде дополнительного яйца в кладке. При этом увеличение вклада самки может проявляться еще и в виде тех положительных эффектов, которые она оказывает на свое потомство.
Следовательно, пара, в которой самец несет аллель сигнализатора, а самка несет аллель респондера (т.е. пары A2B2), имеет дополнительный вклад со стороны самки и, следовательно, более высокую плодовитость, чем другие 3 комбинации.

Варианты комбинаций самцов и самок по соотношению сигналов и ответов на них.
На число потомков, выживших для размножения в следующем году, влияет зависимость от плотности* внутри выводка и зависимость от плотности выводка после оперения.
Зависимость от плотности* — процессы, зависящие от плотности, происходят, когда темпы роста популяции регулируются плотностью этой популяции.
Еще одна группа переменных связана со смертностью самок и самцов после произведения на свет потомства. Эти переменные определяются вкладом в выводок (cm — вклад самцов, cf — вклад самок), затратами на сигналы для самцов (sm) и неселективной смертностью (dm — самцы и df — самки).
Вдовы, вдовцы, несовершеннолетние и любые ранее одинокие особи объединяются, чтобы сформировать новые пары, и годовой цикл завершается. В исследуемой модели акцент ставится на генетической моногамии, потому все типы полового отбора (т.е. конкуренция между особями за партнера) исключаются из расчетов.
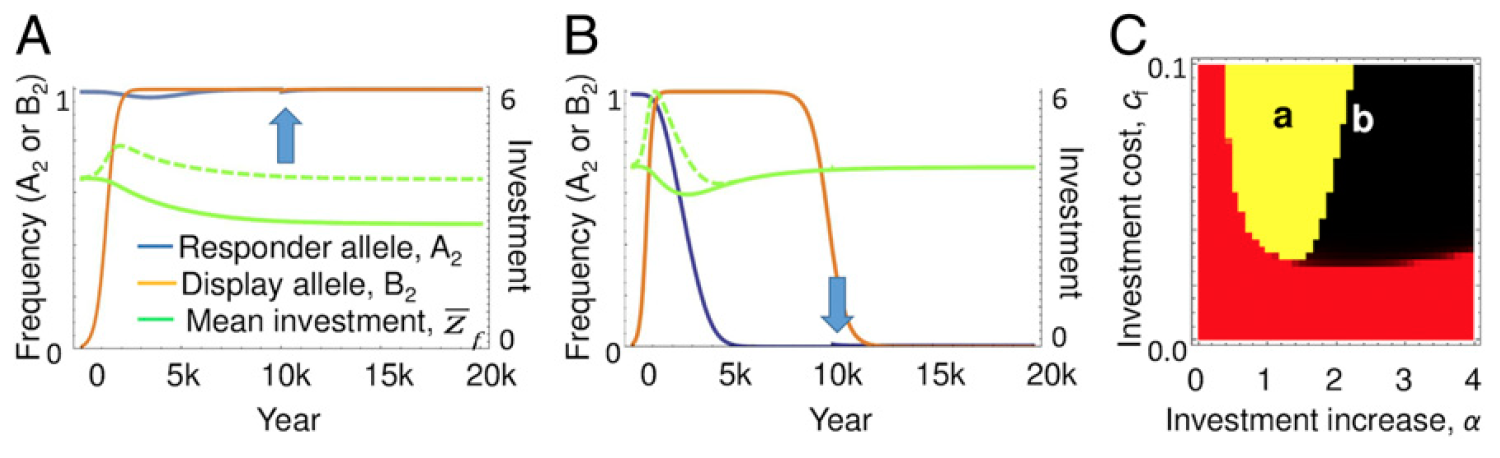
Взаимосвязь эволюции сигналов, респондентов и вклада.
Моделирование показало, что устойчивый баланс достигается, когда самцы подают сигналы, а самки на них отвечают. В состоянии равновесия весь вклад в потомство восстанавливается до того уровня, что был до появления дополнительных сигналов самцов.
На графике А выше показан пример эволюционной динамики, где вклад самок в потомство возвращается к оптимальному уровню, что является результатом эволюции количественного признака вклада (пунктирная зеленая линия — это реальный вклад, а сплошная зеленая линия — это вклад, который не был реализован ввиду отсутствия ответа самок на дополнительные сигналы самцов). На графике В показан альтернативный пример, когда межполовой конфликт приводит к потере респондента.
А на графике С выделены два параметра, которые влияют на такой результат: увеличение вклада, вызванное дополнительными сигналами (α), и затраты самок на этот вклад (cf). В красной области на графике сигналы никогда не увеличиваются, так как их стоимость будет превышать выгоду. В желтой и черной областях частота сигналов возрастает, что приводит к увеличению затратных инвестиций со стороны самок. В желтой области ответ на это происходит путем уменьшения количественного инвестиционного признака, что приводит к постоянной фиксации аллелей как сигналов, так и респондентов. В черной области, где у реагирующих самок больше индуцированных инвестиций, отвечающий аллель быстро теряется, за ним следуют сигналы, как в традиционных моделях межполового конфликта (график В).
Вертикальная граница между красной и желтой областями представляет собой точку, в которой самцы получают дополнительный вклад в потомство за счет того, что самки уравновешивают затраты на проявление ими сигналов. Горизонтальная граница, отделяющая желтую и черную области от красной, возникает аналогично, но по менее очевидной причине. Когда затраты самок на вклад (cf) низкие, то оптимальное значение вклада (zopt) будет относительно высоким, и, следовательно, вклад самок будет значительно больше в начальных условиях. Следствием этого является то, что сигналы дают самцу пропорционально меньшую выгоду от вклада, который он вызывает, что опять же компенсируется его затратами.
Пространство параметров, в котором сигналы и ответы фиксированы (желтый), варьируется в зависимости от силы отбора и генетической дисперсии аллеля респондента. Например, когда начальная частота аллеля респондента составляет 0.9 вместо 0.99, показанной на изображении №2, введение сигналов приводит к более эффективному отбору в отношении респондеров (исходная генетическая дисперсия выше), и черная область расширяется влево.
Сигналы самца могут появиться, даже если они будут сопряжены с затратами, которые уменьшают вклад самца в текущий выводок (параметризованный sfec), тем самым непосредственно влияя на приспособленность* как самца, так и самки, а не уменьшая вероятность выживания самца.
Приспособленность* — способность к размножению особей с определенным генотипом.
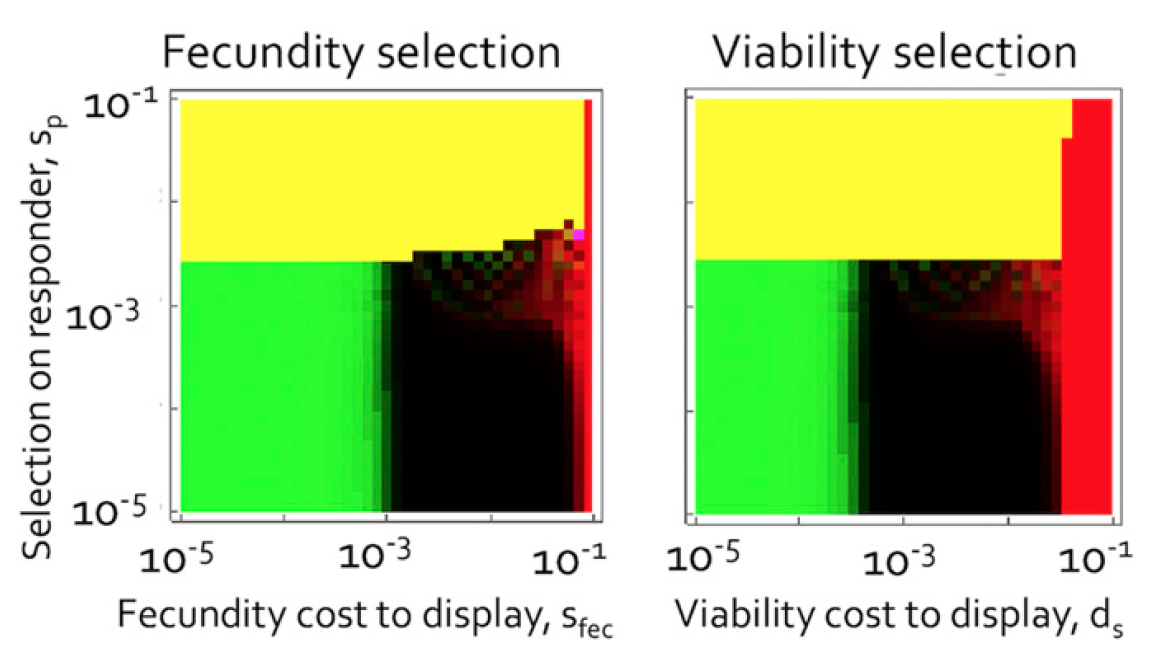
Отношение затрат на плодовитость и сигналов (слева) и отношение затрат на жизнеспособность и сигналов.
В аспекте плодовитости, когда сигналы самца фиксированы (желтая область), все самцы вкладывают меньше в потомство, чем до проявления сигналов. В таком случае вклад самок будет больше, чем он был до проявления сигналов самцов.
Больший вклад самок, когда затраты самцов регулируются плодовитостью (а не жизнеспособностью), увеличивают среднее число потомков на пару, но это не полностью компенсирует. Со временем больший вклад самок приводит к увеличению среднего числа потомства, достигшего оперения, но уменьшает средний показатель жизнеспособности самок. Это приводит к формированию нового баланса между этими двумя силами, где среднее число потомков ниже, чем в случае нормальной жизнеспособности или в начальных условиях (до проявления сигналов).
С математической точки зрения это выглядит следующим образом: если сигналы самцов увеличивают плодовитость на 1% (но не увеличивают жизнеспособность), то затраты самок на потомство увеличиваются на 1.3%, но при этом их смертность также увеличивается на 0.5%, а численность потомства на пару уменьшается на 0.16%.
Если же среднее значение вклада самок изначально ниже оптимального уровня (например, из-за влияния окружающей среды), то при проявлении стимулирующих рост затрат сигналов возникает сбалансированная система, т.е. межполовое сотрудничество. В такой ситуации сигналы самцов не только увеличивают вклад самок в потомство, но и их приспособленность.
Подобное поведение самцов и самок чаще всего возникает ввиду внешних изменений (климат, среда обитания, количество доступной пищи и т.д.). Ввиду этого ученые предполагают, что формирования моногамии у некоторых современных видов, тогда как их предки были полигамными, обусловлено миграцией и, соответственно, сменой окружающей среды.
Для более детального ознакомления с нюансами исследования рекомендую заглянуть в доклад ученых и дополнительные материалы к нему.
Эпилог
Данное исследование продемонстрировало взаимосвязь полигамии и моногамии с точки зрения эволюции. В птичьем царстве самцы всегда старались превзойти друг друга, дабы заполучить внимание самки: ярким оперением, красивым танцем или даже демонстрацией своих строительных способностей. Подобное поведение обусловлено конкуренцией среди самцов, что присуще полигамным видам, чаще всего. С позиции самок все эти сигналы дают возможность оценить качества самца, которые их общее потомство унаследует. Однако с течением времени самцы начали эволюционировать таким образом, чтобы их сигналы были ярче, чем у конкурентов. Самки, в свою очередь, эволюционировали таким образом, чтобы противодействовать подобным сигналам. Ведь всегда должен быть баланс. Если затраты самок на потомство будут несоразмерны с выгодой, то в увеличении затрат нет смысла. Лучше сделать кладку из 3 яиц и выжить в процессе высиживания и воспитания потомства, чем отложить пять и погибнуть, пытаясь их защитить.
Такой межполовой конфликт интересов мог бы привести к катастрофическому падению популяции, но эволюция пошла по более здравому пути — по пути кооперации. В моногамных парах самцы продолжают проявлять себя во всей красе, а самки отвечают на это оптимальным вкладом в потомство.
Любопытно, что мир диких животных не обременен моральными принципами, законами и нормами, а все действия обусловлены эволюцией, генетикой и жаждой продолжения рода.
Возможно для романтиков такое научное объяснение крылатой любви покажется слишком прозаичным, но ученые считают иначе. Ведь что может быть прекраснее, чем эволюционировать именно так, чтобы между самкой и самцом был баланс и настоящее партнерство, учитывающее интересы обеих сторон и нацеленное на благо будущих поколений.
Офф-топ:
Несмотря на то, что название этих птиц не самое красивое («поганковые»), их танец воссоединения просто прекрасен.
Офф-топ 2.0:
Райские птицы являются ярким (буквально) примером того, какое многообразие сигналов самцы посылают самкам в период гнездования (BBC Earth, голос за кадром — Дэвид Аттенборо).
Благодарю за внимание, оставайтесь любопытствующими, ребята! :)

Комментарии
Отправить комментарий