







Интересно о симбиозе
Хотя наше тело - это сплошной сгусток симбиоза, большинство из нас знакомы с этим явлением довольно поверхностно. Пришло время узнать и про столь важную для человека, и вообще жизни на Земле, форму взаимоотношений.

Симбио́з (греч. συμ-βίωσις — «совместная жизнь» от συμ- — совместно + βίος — жизнь) — форма взаимоотношений, при которой оба партнёра извлекают пользу из другого. В природе встречается широкий спектр примеров взаимовыгодного симбиоза (мутуализм). От желудочных и кишечных бактерий, без которых было бы невозможно пищеварение, до растений (примером служат некоторые орхидеи, чью пыльцу может распространять только один, определённый вид насекомых). Такие отношения успешны всегда, когда они увеличивают шансы обоих партнёров на выживание. Осуществляемые в ходе симбиоза действия или производимые вещества являются для партнёров существенными и незаменимыми. В обобщённом понимании такой симбиоз — промежуточное звено между взаимодействием и слиянием.
Часть 1. Возникновение симбиоза.
Типичным примером примитивизации дарвиновских идей является широко распространенное по сей день мнение о том, что в основе эволюции лежат исключительно эгоизм, конкуренция и безжалостная "борьба за существование".
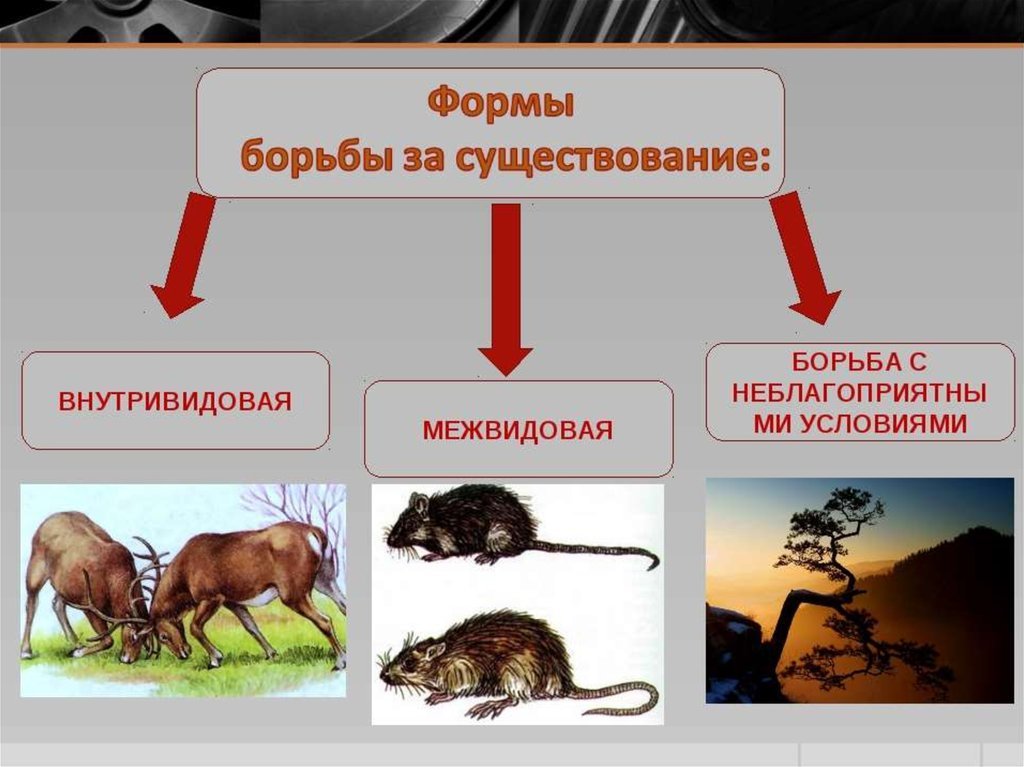
Конкуренция, конечно, играет весьма важную роль и в биологической, и в социальной эволюции. Однако в конечном счете, как правило, в выигрыше оказываются не те, кто сумел добиться безраздельного господства в той или иной сфере и уничтожить всех конкурентов, а те, кому удалось наладить взаимовыгодное сотрудничество с ними и превратить врагов в друзей.

Без сотрудничества, кооперации, симбиоза не может существовать (и тем более развиваться) ни одна живая система. В биологии необходимость кооперации и симбиоза совершенно очевидна. Для того чтобы выжить и оставить потомство, каждое живое существо должно справиться с множеством разнообразных проблем. Нужно каким-то образом получать из окружающей среды необходимые вещества, а недостающие самостоятельно синтезировать из подручного материала; нужно добывать энергию, необходимую для энергоемких химических и физических процессов; нужно вовремя избавляться от отходов жизнедеятельности, находить подходящих партнеров для обмена наследственным материалом, заботиться о потомстве, защищаться от хищников и так далее — и все это в переменчивой, далеко не всегда благоприятной внешней среде. Требования, предъявляемые жизнью к каждому отдельному организму, не только многочисленны и разнообразны — очень часто они еще и противоречивы. Невозможно оптимизировать сложную систему сразу по всем параметрам: чтобы добиться совершенства в чем-то одном, приходится жертвовать другим. Поэтому эволюция — это вечный поиск компромисса, и отсюда следует неизбежная ограниченность возможностей любого отдельно взятого живого существа. Самый простой и эффективный путь преодоления этой ограниченности — симбиоз, то есть кооперация "специалистов разного профиля".
Многие биологи полагают, что все разнообразие жизни на нашей планете происходит от единственного исходного вида — "универсального предка" по имени Лука (Last Universal Common Ancestor, LUCА). Другие, в том числе крупнейший микробиолог академик Г.А. Заварзин, с этим не согласны. Они исходят из того, что устойчивое существование биосферы возможно только при условии относительной замкнутости биогеохимических циклов — в противном случае живые существа очень быстро израсходуют все ресурсы или отравят себя продуктами собственной жизнедеятельности.
Замкнутость циклов может быть обеспечена только сообществом из нескольких разных видов микроорганизмов, разделивших между собой биогеохимические функции. Одни, используя ресурсы среды, наполняют ее продуктами своей жизнедеятельности, а другие, используя эти продукты, возвращают в качестве своих отходов первоначальный ресурс во внешнюю среду. Примером такого сообщества являются циано-бактериальные маты.
Впрочем, следует помнить, что в биологии не бывает правил без исключений и даже видимая безупречность логических рассуждений не всегда может служить критерием истины. Теоретически все-таки можно себе представить единичный вид микроорганизмов, существующий на планете в течение очень долгого времени. Например, если "пищей" ему служат какие-либо вещества, поступающие понемножку из земных недр, а отходы жизнедеятельности либо перерабатываются в геохимических круговоротах без участия живых организмов, либо, к примеру, просто захораниваются в земной коре. Таким образом, этот гипотетический микроорганизм попросту встраивается в уже существующий геохимический цикл, лишь ускоряя его.
Однако в целом на сегодняшний день утверждение "в начале было сообщество" представляется, на мой взгляд, более вероятным, чем "в начале был один вид микробов".
В любом случае, уже 3,55 млрд лет назад на Земле, по-видимому, существовали сложные микробные сообщества — бактериальные маты. В наши дни нечто подобное можно наблюдать в некоторых экстремальных местообитаниях, таких как горячие источники. Древние бактериальные маты, вероятно, состояли из двух слоев. В верхнем обитали аноксигенные (не выделяющие кислород) фототрофы. Они синтезировали органику из углекислого газа, потребляли сероводород и выделяли сульфаты. В нижнем слое жили бродильщики (они потребляли органику, произведенную фототрофами, и выделяли водород), сульфатредукторы (потребляли сульфаты и водород, производили сероводород), а также, возможно, метаногены с метанотрофами.
Важнейшим поворотным пунктом в развитии жизни стало изобретение оксигенного, (кислородного) фотосинтеза, благодаря которому в атмосфере начал накапливаться кислород и стало возможным существование высших организмов. Это великое событие произошло, по-видимому, 2,5-2,7 млрд лет назад (хотя ряд ученых придерживается мнения о более раннем появлении кислородных фотосинтетиков). "Изобретателями" кислородного фотосинтеза были цианобактерии, или, как их раньше называли, сине-зеленые водоросли.

Важность сделанного цианобактериями "открытия" трудно переоценить. Без цианобактерий не было бы и растений, ведь растительная клетка — результат симбиоза нефотосинтезирующего (гетеротрофного) одноклеточного организма с цианобактериями. Все растения осуществляют фотосинтез при помощи особых органелл — пластид, которые суть не что иное, как симбиотические цианобактерии. И неясно еще, кто главный в этом симбиозе. Некоторые биологи говорят, пользуясь метафорическим языком, что растения — всего лишь удобные "домики" для проживания цианобактерий.
После появления цианобактерий безраздельное господство прокариот (безядерных) на нашей планете продолжалось еще очень долго — полтора или два миллиарда лет. Микроорганизмы постепенно становились все более многочисленными и разнообразными. В конце концов уровень интеграции в сообществе прокариот достиг такого уровня, что несколько разных видов микроорганизмов слились в единый организм — эукариотическую клетку.
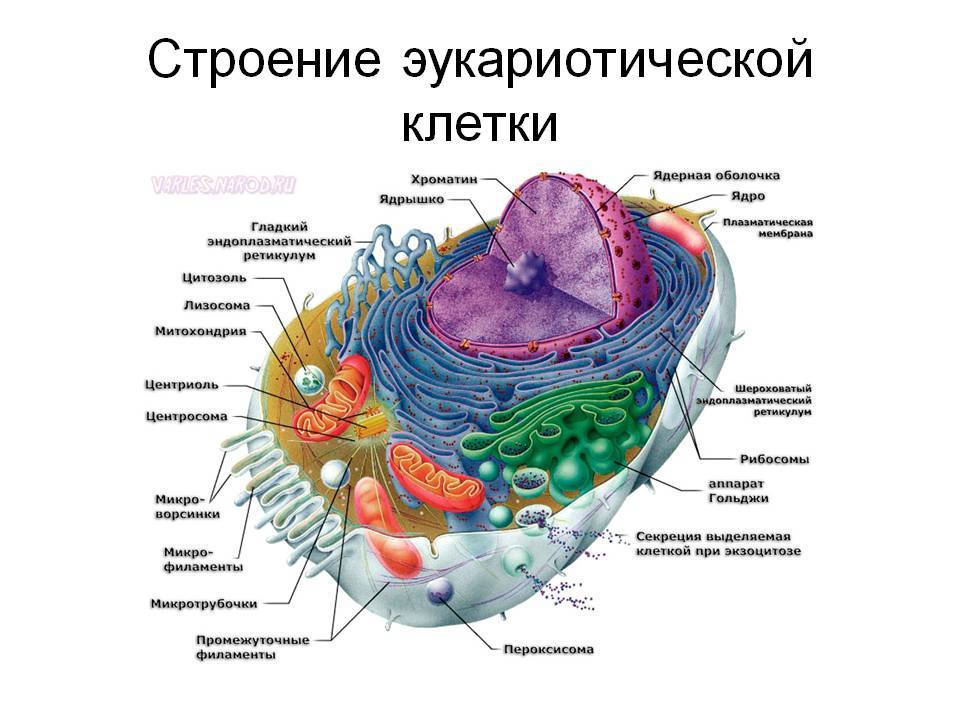
Это событие можно считаеть вторым по значимости в истории жизни на Земле. На первом месте, конечно, стоит само ее появление.
Часть 2. Сообщество превращается в организм.
"Планета микробов" жила и успешно развивалась в течение миллиарда или более лет, и за это время ее микроскопические обитатели добились немалых успехов. Высшим достижением этого этапа эволюции стали сложные микробные сообщества — бактериальные маты.
По уровню целостности бактериальный мат приближается к настоящему организму. Приближается, но все-таки не достигает этого уровня. Прокариоты так и не смогли преодолеть этот рубеж и дать начало настоящим многоклеточным организмам.
Все развитие прокариотной биосферы строилось на основе одного и того же базового "кирпичика" - прокариотической клетке. При всех бесспорных преимуществах этой универсальной живой единицы, при всем ее умении приспосабливаться почти к любым условиям и извлекать энергию чуть ли не из любого химического процесса в строении прокариотической клетки есть ряд непреодолимых ограничений.
Главное из них связано с отсутствием клеточного ядра. Наследственный материал прокариот (обычно это единственная кольцевая молекула ДНК — кольцевая хромосома) находится прямо в цитоплазме, то есть, образно говоря, в бурлящем биохимическом котле, где происходит обмен веществ и осуществляются тысячи химических реакций. В такой неспокойной обстановке очень трудно развить сложные и эффективные молекулярные механизмы регуляции работы генов.
Эукариоты же, спрятав ДНК в ядре, получили недосягаемую прежде возможность тонкой регуляции работы генов.

Это позволило клетке радикально менять свои свойства, структуру и облик, не изменяя при этом сам геном, а только усиливая или ослабляя работу разных генов. Именно эта пластичность позволила эукариотам в конце концов стать многоклеточными в строгом смысле этого слова. Ведь в настоящем многоклеточном организме не просто много клеток, а много разных типов клеток (покровные, мышечные, нервные, половые и т. д.) Однако геном у них у всех один и тот же!

Второй важный конструктивный "недостаток" прокариотической клетки состоит в отсутствии окруженных мембранами органелл. А ведь далеко не все биохимические процессы, которые могли бы оказаться полезными клетке, можно осуществлять в едином "общем котле", каковым является цитоплазма прокариот. Представьте себе, как усложнилась бы работа химика, если бы в его распоряжении имелась лишь одна-единственная пробирка! Прокариоты, конечно, пытались по-своему преодолеть этот недостаток. Если присмотреться к прокариотической клетке повнимательнее, можно заметить, что в распоряжении бактерий на самом деле не одна "пробирка", а две. В роли второй область снаружи от клеточной мембраны (здесь обычно расположено толстое, рыхлое, сложно устроенное многослойное образование — "клеточная стенка"). В толще клеточной стенки могут протекать химические процессы, не совместимые с теми, которые идут в цитоплазме. Но и двух пробирок все-таки мало для хорошей химической лаборатории!

Эукариоты в этом отношении дадут прокариотам сто очков вперед. Внутренняя среда эукариотической клетки подразделена двойными и одинарными мембранами на множество разнообразных отсеков (ядро, митохондрии, пластиды, эндоплазматическая сеть и т. д.).
Именно из-за этих ограничений прокариоты с самого начала так сильно тяготели к симбиозу. В микробном сообществе разные виды микробов в функциональном отношении соответствуют разным отсекам эукариотической клетки. Сообща микробы могут достичь гораздо большего, чем поодиночке.

Однако как бы ни были тесны связи в симбиотическом содружестве микробов, сообщество не становится настоящим целостным организмом, потому что оно не может размножаться как единое целое. Каждый из микробов сохраняет способность "сбежать" из сообщества и перейти к самостоятельной жизни, если где-то сложатся подходящие для этого условия; каждый размножается сам по себе и имеет свой собственный индивидуальный геном, который и передает своим потомкам. Поэтому естественный отбор продолжает действовать на уровне отдельных клеток, а не на уровне сообщества. Любая мутация, повышающая жизнеспособность данного конкретного микроба, будет поддержана отбором даже в том случае, если это вредно для сообщества как целого.

Чтобы преодолеть эти ограничения, прокариотическим клеткам нужно было сделать еще один шаг — вполне естественный и логичный — в сторону дальнейшего усиления интеграции, сплоченности сообщества. Они должны были по-настоящему слиться в единый организм, отказаться от своей клеточной индивидуальности и объединить свои персональные хромосомы в один большой общий геном.
Именно это и случилось в начале протерозойского эона (около 2,0-2,2 млрд лет назад)
В настоящее время в научной литературе обсуждается два или три десятка возможных сценариев превращения сообщества прокариот в эукариотическую клетку. Они разнятся в деталях, но имеют много общего в целом. Общепризнанным считаются следующее: митохондрии и пластиды являются прямыми потомками прокариот. В качестве неоспоримых доказательств своего бактериального происхождения они по сей день сохранили кольцевые хромосомы и способность к самостоятельному размножению путем деления. То есть, даже по прошествии двух с лишним миллиардов лет они все еще держат свои чемоданы собранными.

Часть 3. Как бактерии превращаются в органеллы
После приобретения митохондрий и пластид эукариоты вовсе не утратили способности к заглатыванию и "приручению" бактерий. Этот процесс продолжается и по сей день. Многие одноклеточные эукариоты (амебы, инфузории и другие) прямо-таки нашпигованы всевозможными симбиотическими прокариотами. Например, инфузории, обитающие в рубце жвачных, и жгутиконосцы, населяющие кишечник термитов, содержат в своей цитоплазме симбиотических бактерий, помогающих им переваривать клетчатку (целлюлозу). Такие симбиотические системы напоминают матрешку: в корове — инфузории, в инфузориях — бактерии.

Зачем корова так тщательно пережевывает траву? Очень просто: кусочки травы должны стать достаточно маленькими, чтобы их могли проглотить симбиотические инфузории. Но и инфузория сама не может переварить целлюлозу и перепоручает это непростое дело своим собственным симбионтам — бактериям.
Иногда бактерии и сами проявляют инициативу, изобретая различные способы проникновения в клетки эукариот. В этом случае велика вероятность того, что внутриклеточная бактерия станет не полезным симбионтом, а вредным паразитом. Впрочем, независимо от того, как будут складываться взаимоотношения внутриклеточных бактерий с хозяином, судьба их в некотором смысле предопределена.

Жизнь внутри чужой клетки способствует постепенному упрощению и деградации. Внутриклеточные бактерии начинают идти по тому же пути, по которому когда-то прошли митохондрии и пластиды, то есть теряют свои гены, становятся все более зависимыми от хозяина и постепенно превращаются из самостоятельных организмов в нечто, очень напоминающее органеллы. У одних внутриклеточных бактерий этот процесс зашел уже очень далеко, у других он только начинается.

Мы рассмотрим два самых крайних случая, в одном из которых внуктриклеточная бактерия, можно сказать, еще даже не начала упрощаться и терять гены, а в другом — практически уже стала органеллой, побив все рекорды генетической дегенерации.
Первая из двух рекордсменок — бактерия Ruthia magnified, внутриклеточный симбионт, обитающий в тканях двустворчатого моллюска Calyptogena magnifica.
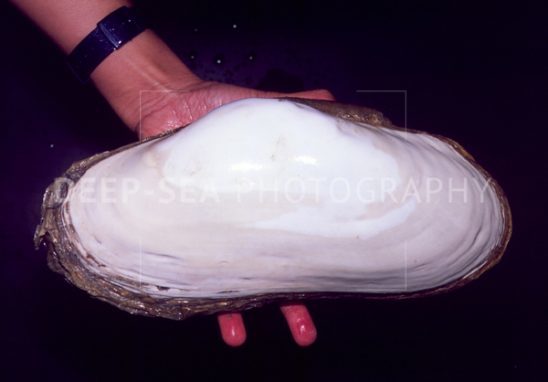
Этот симбиотический "сверхорганизм" обитает на дне моря на большой глубине вблизи гидротермальных источников. Надо сказать, что дно океанов на глубине нескольких километров обычно довольно пустынно, и вовсе не потому, что живые организмы не выдерживают высокого давления. Лимитирующим фактором является пища: свет сюда не проникает, фотосинтез невозможен, и донные животные могут питаться лишь теми скудными крохами, которые падают сверху, из освещенных слоев воды, где жизнь гораздо богаче.
Однако там, где из морского дна просачивается сероводород или метан, расцветают настоящие оазисы жизни. Существуют они за счет хемоавтотрофных бактерий, окисляющих H2S и СН4 при помощи кислорода.
Тут я прошу на минуту отвлечься и осознать: они едят сероводород и метан!

Выделяющуюся в ходе этой химической реакции энергию бактерии используют для синтеза органических веществ из углекислого газа. Все животные, обитающие в гидротермальных оазисах (черви, моллюски, креветки), либо питаются этими бактериями, отфильтровывая их из воды, либо вступают с ними в симбиоз, поселяя бактерий на поверхности своего тела или даже внутри него. Сообщества морских гидротерм интересны тем, что они, в отличие от большинства других экосистем, существуют за счет энергии земных недр, а не солнечного света. Они почти независимы от окружающей биосферы, если не считать того, что кислород, образуемый где-то там наверху фотосинтезирующими организмами, им все-таки необходим.

Автотрофные внутриклеточные симбионты гидротермальных животных по своей функциональной роли напоминают пластиды. Разница лишь в источнике энергии, используемой для синтеза органики: пластиды используют солнечный свет (то есть являются фотоавтотрофами), а гидротермальные бактерии — энергию химических реакций (хемоавтотрофы).
Гигантский моллюск Calyptogena magnifica был одним из первых гидротермальных животных, описанных в научной литературе (его описали в 1980 году, вскоре после открытия глубоководных гидротерм). Пищеварительная система моллюска сильно редуцирована, и почти все необходимое он получает от живущего в его клетках бактериального симбионта.
Как выяснилось, у Ruthia magnifica, несмотря на внутриклеточный образ жизни, генетическая дегенерация зашла совсем недалеко. Размер генома бактерии — 1,2 млн пар нуклеотидов. Для свободноживущих бактерий это маловато, но для внутриклеточных — очень много (больше, чем у всех прочих внутриклеточных бактерий с прочтенным геномом). У Ruthia magnifica, судя по набору генов, сохранились все метаболические пути, характерные для свободноживущих хемоавтотрофов. У нее присутствуют, например, все гены, необходимые для фиксации С02, для окисления восстановленных соединений серы, для синтеза различных кофакторов и витаминов, а также всех 20 аминокислот (в этом отношении Ruthia превосходит всех остальных изученных внутриклеточных микробов).
Анализ генома подтвердил, что бактерия, как и предполагалось, фиксирует неорганический углерод при помощи несколько искаженного цикла Кальвина, а энергию для этого получает за счет окисления соединений серы. Когда сероводород имеется в избытке, бактерия окисляет его до серы, которая в виде гранул хранится в бактериальных клетках "на черный день". Эти запасы впоследствии могут использоваться для дальнейшего окисления (до сульфита, а затем и до сульфата, который выводится из клетки специальными белками-транспортерами). В геноме бактерии имеются все необходимые гены для осуществления этих реакций.
Моллюск-хозяин активно снабжает своего симбионта необходимой ему пищей: в крови моллюска обнаружен особый цинк-содержащий белок, предназначенный для связывания и транспортировки сероводорода. У бактерии есть также полный набор генов, необходимых для кислородного дыхания. Это означает, что микроб, как и предполагалось, использует в качестве окислителя кислород, которым его тоже обеспечивает хозяин. Имеются также наборы генов для таких важных метаболических путей, как гликолиз и цикл Кребса. Все это есть и у хозяина, и бактерия могла бы просто брать готовые продукты извне, однако не делает этого.

Бактерия снабжает своего хозяина аминокислотами и витаминами. Кроме того, она способна утилизировать отходы его жизнедеятельности, такие как аммоний. Это вещество бактерия всасывает из тканей хозяина и использует к обоюдной пользе, например, для синтеза тех же аминокислот.
Главной особенностью Ruthia magnifica, конечно, является ее удивительная биохимическая самостоятельность. Такого полного набора генов, необходимых для основных биохимических процессов, свойственных свободноживущим хемоавтотрофам, ни у одной другой внутриклеточной бактерии не обнаружено. По-видимому, симбиотическая система Calyptogena magnifica — Ruthia magnifica находится на ранней стадии эволюционного становления и симбионт еще не успел далеко продвинуться по пути неизбежной в его положении деградации.
Противоположный пример, показывающий, как далеко может зайти микроб по пути превращения в органеллу, дает бактерия Carsonella. Карсонелла живет в клетках листоблошек — мелких, похожих на тлей насекомых, питающихся исключительно соком растений.

Как и другие насекомые, придерживающиеся этой более чем скромной диеты (например, тли и клопы), листоблошки обзавелись бактериальными помощниками, которые синтезируют для них необходимые вещества, отсутствующие в растительном соке, в первую очередь аминокислоты. Тут вернее будет сказать, что удачный симбиоз оказался решающим фактором, который позволил листоблошкам (и другим насекомым) перейти на питание чистым растительным соком. Предполагается, что перед тем, как стать внутриклеточными симбионтами, предки карсонеллы жили в кишечнике насекомых.
У карсонеллы наблюдаются все три основных признака генетической деградации, свойственной внутриклеточным бактериям: сокращение генома в результате потери почти всех некодирующих участков ДНК и значительной части генов, резкое преобладание в ДНК нуклеотидов А и Т и, соответственно, низкое содержание Г и Ц и быстрая молекулярная эволюция, то есть повышенный темп изменения ДНК в ряду поколений.
По первому и второму пункту карсонелла побила все прежние рекорды. Ее геном втрое меньше, чем у архей Nanoarchaeum equitans, которая живет в гидротермальных источниках и паразитирует на другой архее — Ignicoccus (Это единственный случай, когда один прокариотический организм является облигатным (обязательным) паразитом другого)— и одной из разновидностей бактерии Buchnera, внутриклеточного симбионта тлей. У этих двух прокариот размер генома составляет 450-490 тысяч пар нуклеотидов (т.п.н.), и раньше именно они считались рекордсменами генетического упрощения. Карсонелла с ее 160 т.п.н. оставляет конкурентов далеко позади.

Геном карсонеллы по размеру вполне сопоставим с геномом митохондрий. Типичные размеры митохондриальных геномов: 40-100 т.п.н. у низших эукариот, 200-400 т.п.н. у растений, 15-20 т.п.н. у животных.
Само собой разумеется, что карсонелла не может жить вне клеток хозяина и передается только вертикально — от матери к ее детям (как и митохондрии). Сохранившихся у карсонеллы генов явно недостаточно для поддержания ее жизни — даже с учетом того, что она может пользоваться всеми благами внутриклеточного существования. Очевидно, что специализированные хозяйские клетки — бактериоциты — целенаправленно поддерживают жизнь симбионтов. Вполне возможно (хотя и не доказано наверняка), что многие гены, утраченные предками карсонеллы, были перенесены в геном хозяина, где они продолжают функционировать, обеспечивая бактерию необходимыми веществами извне. Как мы помним, именно это произошло когда-то с генами предков митохондрий.
Часть 4. Вездесущий симбиоз.
Когда-то симбиоз считался сравнительно редким явлением — скорее курьезом, чем правилом. Когда в 60-е годы XIX века было обнаружено, что лишайники представляют собой симбиотические комплексы из грибов и водорослей, ученые страшно удивились (надо же, какие причуды бывают у матушки-природы!).
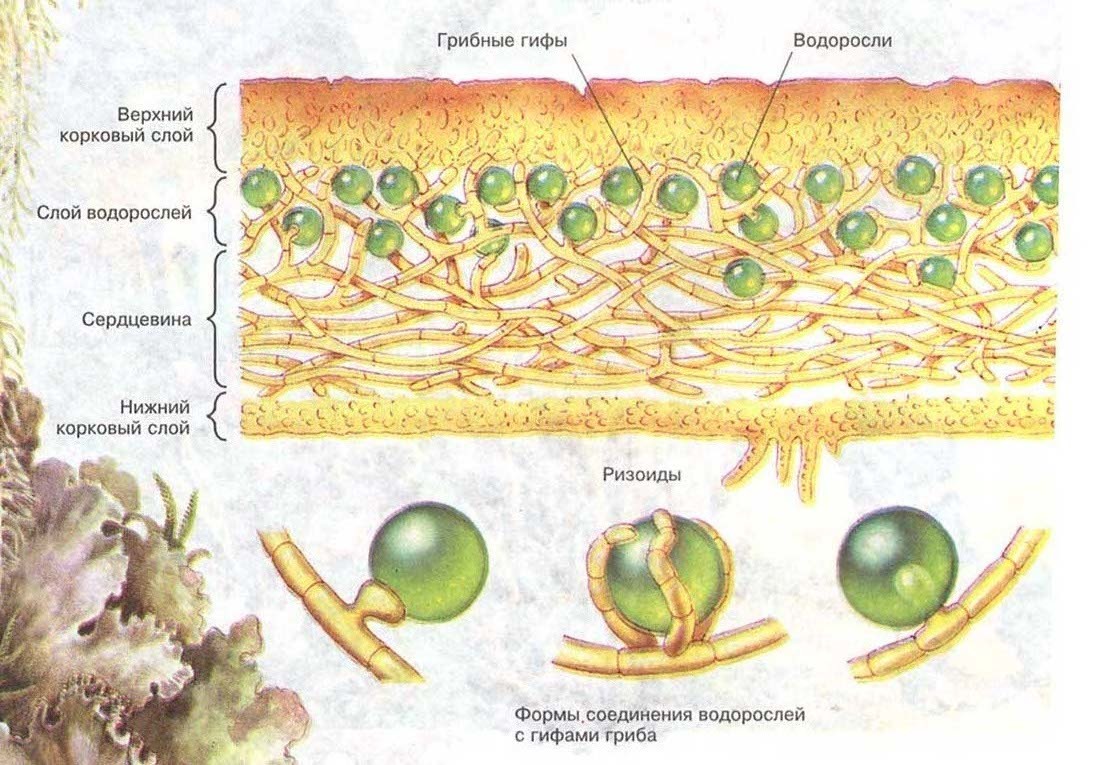
С тех пор многое изменилось. Уже в начале XX века отдельные выдающиеся мыслители предполагали, что симбиоз и кооперация могут играть огромную роль в развитии жизни на Земле. Хотя "организмоцентрический" подход в биологии по-прежнему господствует, сегодня ученые ясно понимают, что симбиоз — это магистральный путь эволюции, без которого прогрессивное развитие жизни на Земле было бы крайне затруднено, если вообще возможно.
По-настоящему "автономный организм", сформировавшийся и живущий без всякого участия каких-либо симбионтов, в природе еще надо поискать. Большинство живых существ, населяющих планету, в действительности являются "сверхорганизмами" — сложными симбиотическими комплексами.
Человек не исключение. Каждая наша клетка получает необходимую ей энергию от митохондрий — потомков симбиотических бактерий. Многие из наших генов получены нами от вирусов, всевозможных "эгоистических" фрагментов ДНК и мобильных генетических элементов. Наш метаболизм во многом определяется многочисленными микробами, составляющими кишечную флору. И даже если заглянуть внутрь любого из этих микробов, то и там мы найдем сожителей-симбионтов (плазмиды, фаги, транспозоны).
Кишечная микрофлора превращает человека в "сверхорганизм". По имеющимся оценкам, в кишечнике взрослого человека присутствует более 1 кг микроорганизмов, относящихся к сотням различных видов. В точности их видовой состав неизвестен. Микробиологи знают "в лицо" лишь несколько десятков типичных представителей, которых можно вырастить на искусственных средах. Как выяснилось сравнительно недавно (и это открытие стало шоком для микробиологов), большинство существующих в природе микроорганизмов на искусственных средах не растет.

В "явном виде", применив метагеномный анализ, по генам рибосомной РНК удалось идентифицировать лишь 72 разновидности бактерий (из них 60 некультивируемых и 16 новых для науки) и один вид архей-метаногенов, однако статистически обосновано число микробов составляет не менее 300.

На основе проведенного анализа были определены наиболее важные метаболические функции, которые выполняют микробы в человеческом кишечнике. Это прежде всего переваривание растительных полисахаридов, которые не могут перевариваться ферментами, закодированными в геноме человека. С этими трудноусваиваемыми углеводами расправляются в основном бактерии-бродильщики, выделяющие в качестве конечных продуктов обмена низкомолекулярные органические кислоты. Однако то, что для бактерий-бродильщиков является отходами жизнедеятельности, для человека — вполне съедобные вещества, которые активно всасываются кишечным эпителием. По имеющимся оценкам, из этого необычного источника люди получают около 10% калорий (эта оценка справедлива для приверженцев типичной "европейской" диеты). Задумайтесь, что это значит: не бактерии питаются нашими отходами, как кто-то мог бы подумать, а как раз наоборот!

Кроме съедобных для человека веществ бактерии-бродильщики выделяют в качестве побочного продукта еще и молекулярный водород, который вреден для них самих и препятствует их росту и жизнедеятельности. Чтобы процесс переваривания растительных полисахаридов шел эффективно, кто-то должен постоянно утилизировать образующийся водород. Именно этим и занимаются археи-метаногены. В ходе метаногенеза поглощаются водород и углекислый газ и выделяется метан.
В "совокупном геноме" кишечной флоры сильно повышено процентное содержание генов, связанных с синтезом незаменимых аминокислот и витаминов. Микробы сильно облегчают человеку жизнь, производя значительные количества этих необходимых нам веществ. Кроме того, кишечная флора располагает большим арсеналом ферментов для обезвреживания токсичных веществ, присутствующих в нашей повседневной пище, особенно растительной.
С учетом инеющихся даных человека следует рассматривать как "сверхорганизм", чей обмен веществ обеспечивается совместной слаженной работой ферментов, закодированных не только в геноме Homo sapiens, но и в геномах сотен видов симбиотических микробов. Причем, доля человеческих генов в совокупном геноме этого "сверхорганизма" составляет не более 1%.

На симбиозе были основаны многие важнейшие ароморфозы (прогрессивные эволюционные преобразования). Мы уже познакомились с самым значительным из них — формированием эукариотической клетки, той основы, из которой в дальнейшем развились все высшие формы жизни. В дальнейшем роль симбиоза в развитии жизни отнюдь не снижалась. Важнейшие функциональные блоки современной биосферы целиком и полностью держатся на симбиозе и симбиотических комплексах — "сверхорганизмах".
Так, возможности высших растений — основных производителей органики и кислорода — были бы весьма ограничены без симбиоза с бактериями, способными переводить атмосферный азот в доступную для растений форму, и с почвенными грибами (микориза), без кооперации с насекомыми-опылителями и позвоночными — распространителями семян.


Растительноядные животные — основные потребители производимой растениями органики — не могут эффективно переваривать растительную пищу без помощи разнообразных симбиотических бактерий и одноклеточных эукариот.
Самые яркие и богатые жизнью морские экосистемы коралловых рифов невозможны без симбиоза коралловых полипов с одноклеточными водорослями — зооксантеллами.
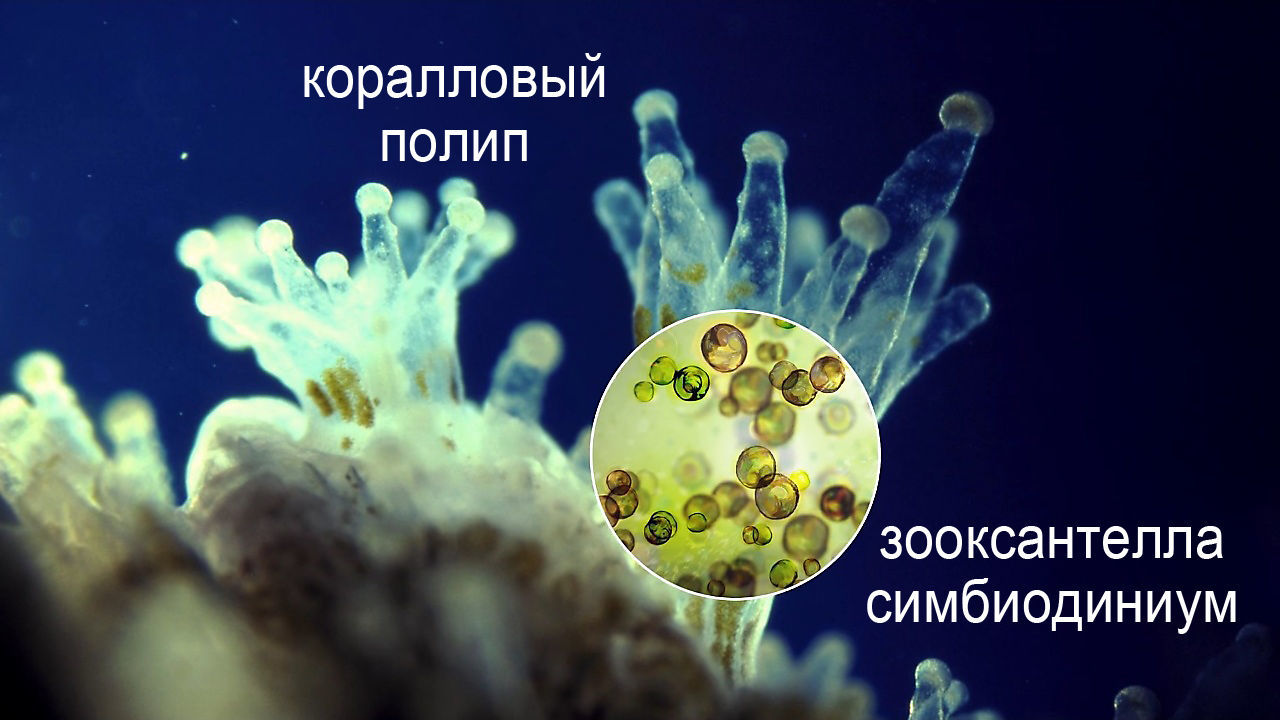
Сообщества различных экзотических, архаичных и экстремальных местообитаний (таких как наземные и подводные горячие источники, выходы метана и сероводорода, соленые лагуны, подземные воды и др.) тоже сплошь и рядом представляют собой сложные симбиотические комплексы микроорганизмов, в которых порой принимают участие и высшие организмы.

Выход растений на сушу — результат симбиоза. Первые наземные растения появились в конце силурийского периода (более 400 млн лет назад). Недавно палеонтологи обнаружили, что уже самые первые наземные растения жили в симбиозе с грибами: у них была самая настоящая микориза. У этих растений еще не было настоящих корней — вместо них имелись так называемые ризоиды, не способные самостоятельно всасывать что-либо из почвы и служившие только для закрепления в грунте, а также, как теперь выяснилось, для обеспечения симбиоза с почвенными грибами. По- видимому, без этого симбиоза растения вообще не смогли бы выйти на сушу. Симбиоз растений с азотфиксирующими бактериями возник позже, причем для этого растениям оказалось достаточно лишь немного изменить те генетические системы, которые сложились у них ранее для взаимодействия с микоризными грибами.

Симбиоз с автотрофами открывает большие возможности для многих водных животных, особенно малоподвижных (кишечнополостных, губок, асцидий, некоторых червей и моллюсков). Такие симбиотические комплексы представляют собой "сверхорганизмы", сочетающие признаки растений и животных (яркий пример — коралловые полипы). Автотрофы не только снабжают хозяина органикой, полученной в результате фото- или хемосинтеза, но и в ряде случаев помогают ему избавляться от конечных продуктов азотного обмена (например, мочевой кислоты или мочевины), которые служат для симбионтов ценным источником азота.

Совместное существование позволяет каждому из компонентов симбиотического комплекса отказаться от тех биохимических функций, которые лучше удаются партнеру, и сосредоточиться на том, что лучше получается у него самого. Например, клубеньковые бактерии занимаются почти исключительно фиксацией азота, переложив заботу обо всем остальном на растение-хозяина. Часто один из партнеров специализируется на поставке в систему азота, а другой — углерода. По мере развития симбиотических отношений симбионт может полностью утратить самостоятельность и превратиться в неотъемлемую часть своего хозяина (впрочем, надо признать, что и хозяин при этом теряет самостоятельность и перестает быть "автономным организмом").
Грань между мутуалистическими (взаимовыгодными) и антагонистическими симбиозами — довольно условная и зыбкая. Например, многие растительно-грибные симбиозы в ходе эволюции могли долго "балансировать" на грани мутуализма и антагонизма, причем преобладающие потоки питательных веществ могли неоднократно менять свое направление. Непосредственный переход паразитизма в мутуализм — сранительно редкое явление (пример — "защитные симбиозы" растений со спорыньевыми грибами, в которых исходно паразитический гриб стал защищать растение от растительноядных животных путем синтеза токсичных веществ). В большинстве случаев симбиотические системы развиваются из фрагментов сообществ (экосистем). Например, симбиозы, основанные на биохимической кооперации, в большинстве случаев развиваются из кооперативных объединений свободноживущих организмов, соместно утилизирующих какой-то ресурс, или из фрагментов "трофической пирамиды" (симбиозы производителей органики с ее потребителями). Особый и весьма удивительный случай связи между трофической цепью и биохимическим симбиозом представляет морской моллюск Elysia viridis, питающийся водорослями. Этот моллюск ухитряется переселять пластиды съеденных водорослей в свои собственные клетки и долгое время сохранять их там живыми, приобретая таким образом способность к фотосинтезу. Настоящий гибрид животного и растения.

Часть 5. Экстремальный симбиоз
Термостойкая трава
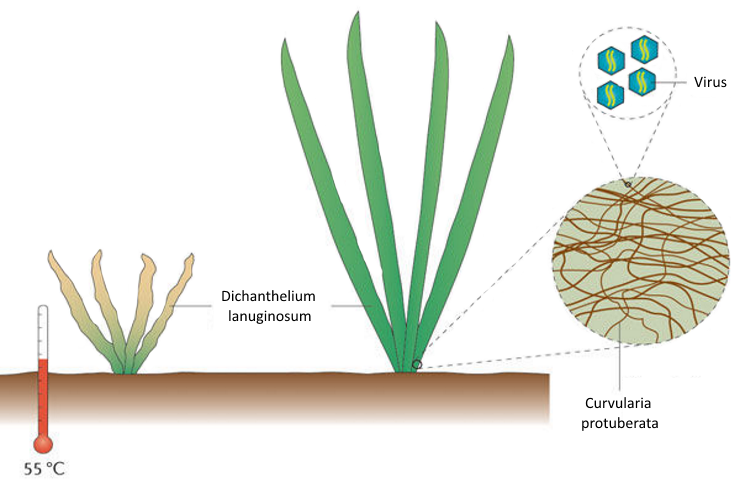
Фантастический случай тройного симбиоза описали в начале 2007 года американские биологи, работающие в Иеллоустонском национальном парке (США), где на горячей почве вблизи геотермальных источников произрастает термостойкая трава Dichanthelium lanuginosum, близкая родственница проса.

Ранее было установлено, что удивительная устойчивость этого растения к высоким температурам каким-то образом связана с произрастающим в тканях растения грибом Curvularia protuberata. Если выращивать растение и гриб по отдельности друг от друга, ни тот ни другой организм не выдерживает длительного нагревания свыше 38°С, однако вместе они прекрасно растут на почве с температурой 65°С. Кроме того, даже в отсутствие теплового стресса растение, зараженное грибом, растет быстрее и лучше переносит засухи.
Продолжая исследование этой симбиотической системы, ученые обнаружили, что в ней есть еще и третий обязательный участник — РНК-содержащий вирус, обитающий в клетках гриба.
Оказалось, что гриб, "вылеченный" от вируса, не в состоянии сделать растение термоустойчивым. Растения с таким грибом погибали на горячей почве точно так же, как и растения без гриба.
Проведенные впоследствии эксперименты на томатах показали, что гриб, зараженный вирусом, способен повышать термоустойчивость не только у своего природного хозяина — однодольного растения Dichanthelium lanuginosum, но и у неродственных растений, относящихся к классу двудольных.
Вместо выделительной системы — микробное сообщество
Еще более фантастическим случаем симбиоза является малощетинковый кольчатый червь Olavius algarvensis, обитающий в Средиземном море. Червь этот интересен прежде всего тем, что у нет ни рта, ни кишечника, ни ануса, ни нефридиев — органов пищеварения и выделения. Некоторые другие морские черви тоже научились обходиться без органов пищеварения. Например, у погонофор кишечник превратился так называемую трофосому — тяж, набитый симбиотическими бактериями, окисляющими сероводород или метан. Поэтому можно было ожидать, что и у Olavius algarvensis отсутствие кишечника компенсируется наличием каких-то симбиотических микробов, обеспечивающих своего хозяина пищей в обмен на беззаботную жизнь в чужом теле. Однако редукция еще и выделительной системы — это явление беспрецедентное для кольчатых червей.

Метагеномный анализ выявил присутствие в теле червя четырех видов симбиотических бактерий, два из которых относятся к группе гамма-протеобактерий, а два других — к дельта-протеобактериям. Обе гамма-протеобактерии, геном которых удалось ренконструировать почти полностью, являются автотрофами, то есть синтезируют органические вещества из углекислого газа. Необходимую для этого энергию они получают за счет окисления сульфида (S2-). В качестве окислителя используется кислород, а при отсутствии кислорода — нитраты (Использование нитратов в качестве окислителя для получения энергии называют нитратным дыханием. Нитратное дыхание широко распространено у бактерий, а недавно его обнаружили и у одноклеточных эукариот - фораминифер, родственников амеб). Если же нет под рукой и нитратов, окислителем могут служить некоторые органические вещества. В качестве конечных продуктов жизнедеятельности эти бактерии выделяют окисленные соединения серы (например, сульфаты).

Погонофоры и вестиментиферы ранее считались отдельным типом животных, но впоследствии было показано, что они являются сильно видоизмененной группой кольчатых червей. Сейчас их называют сибоглинидами. Они встречаются в больших количествах возле подводных гидротермальных источников, где много метана или сероводорода. Сибоглиниды являются по сути дела автотрофными животными. Если быть совсем точным, это автотрофные симбиотические "сверхорганизмы". Живущие в их трофосоме (бывшем кишечнике) хемоавтотрофные бактерии синтезируют органику из углекислого газа, а энергию для этого получают из химической реакции окисления сероводорода или метана. Червь питается органикой, производимой симбиотическими бактериями. Кровь червя переносит не только кислород, необходимый как червю, так и бактериям, но и сероводород — "пищу" бактерий.

Дельта-протеобактерии тоже оказались автотрофами, но другого рода, а именно сульфат-редукторами. Они получают энергию, восстанавливая сульфат (или другие окисленные соединения серы) до сульфида. Таким образом, метаболизм гамма- и дельта-протеобактериальных симбионтов оказался взаимодополнительным: отходы первых служат пищей вторым и наоборот.
В качестве восстановителя (донора электронов, необходимого для восстановления сульфата) симбиотические дельта-протеобактерии могут использовать молекулярный водород. У них есть гены ферментов — гидрогеназ, необходимых для работы с молекулярным водородом. Возможно (хотя и не удалось доказать наверняка), что гамма-протеобактериальные симбионты производят некоторое количество Н2 и таким образом снабжают дельта-протеобактерий не только окисленными соединениями серы, но и восстановителем.
Бактериальные симбионты живут не в глубине тела, а прямо под наружной оболочкой (кутикулой) червя. Здесь они ведут свою странную микробную жизнь, обмениваясь друг с другом продуктами своего метаболизма. Все прочее, чего им может недоставать, они получают из окружающей среды — в основном это вещества, просачивающиеся из морской воды под кутикулу хозяина. Микробы размножаются, а эпителиальные клетки червя тем временем потихоньку заглатывают их и переваривают. Этого источника питания, очевидно, червю вполне достаточно, чтобы не испытывать дискомфорта из-за отсутствия рта и кишечника.

Но как удается червю обходиться без выделительной системы? Оказалось, что в геномах бактерий-симбионтов присутствуют гены белков, обеспечивающих всасывание и утилизацию мочевины, аммония и других отходов жизнедеятельности червя. Эти вещества служат бактериям ценными источниками азота.
Возможно, основная выгода, которую бактерии получают от сожительства с червем, состоит в том, что он подвижен и может по мере надобности переползать туда, где условия среды наиболее благоприятны для всей честной компании. В верхних слоях осадка, где имеется немного кислорода, но нет сульфидов, гамма-протеобактерии могут получать необходимые им сульфиды от своих сожителей — дельта-протеобактерий. Сульфид в этом случае будет окисляться кислородом — наиболее энергетически выгодным окислителем. В больших количествах, правда, кислород вреден для сульфат-редукторов — дельта-протеобактерий.
Если червь закопается поглубже, он попадет в слои, где кислорода нет вовсе. Здесь гамма-протеобактерии будут использовать в качестве окислителя нитраты, что несколько менее выгодно, зато сульфида у них будет вдоволь, потому что кислород больше не будет угнетать жизнедеятельность дельта-протеобактерий.
Наконец, в еще более глубоких слоях осадка, где нет не только кислорода, но и нитратов, гамма-протеобактерии могут использовать в качестве окислителя некоторые органические вещества, выделяемые червем-хозяином и дельта-протеобактериями. При этом в клетках одного из двух видов гамма-протеобактерий запасается сера (как продукт неполного окисления сульфида), которую можно до- окислить позже, когда червь выползет повыше и станут доступны более сильные окислители. Щавелевая кислота, выделяемая гамма-протеобактериями, охотно утилизируется дельта-протеобактериями, и так далее: исследователи выявили еще целый ряд возможных способов "биохимического сотрудничества" внутри этого удивительного симбиотического комплекса.
Таким образом, пять видов живых существ, объединившись, превратились в универсальный "сверхорганизм", способный жить в самых разнообразных условиях — в том числе и там, где ни один из его "компонентов" не выжил бы в одиночку.

А. Марков "Рождение сложности"

Комментарии
О сколько нам обломов в жизни готовит просвещенья дух!
О сколько нам обломов в жизни готовит просвещенья дух!
Отправить комментарий